Меню
Меню





 Все темы
Все темы
Высокогорные карповые рыбы становятся менее полиплоидными
Считается, что предки всех позвоночных животных прошли через этап аутополиплоидизаци, после которого вернулись к диплоидному геному. Однако детали этого предполагаемого события неизвестны. Авторы статьи в Nature получили de novo геномы обитающих в водоемах Тибетского плато карповых рыб подсемейства Schizothoracine — все его представители полиплоидны. Проанализировав структуру их генома, ученые пришли к выводу, что общий предок Schizothoracine стал тетраплоидным из-за удвоения всего генома, а затем из-за слияния отдельных квартетов хромосом начал возвращаться к диплоидному состоянию. Однако на момент эволюционного расхождения видов диплоидизация прошла не полностью и, по-видимому, продолжается до сих пор, из-за чего у одной из этих рыб — Schizopygopsis younghusbandi — тетраплоидный набор содержит 90 хромосом, хотя гаплоидный геном состоит из 25.
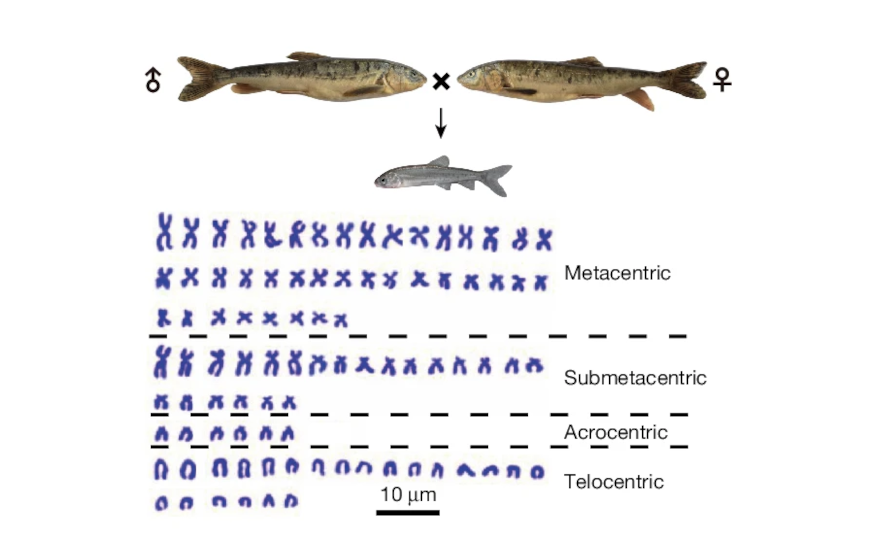
Трио «родитель-потомок» вида S. younghusbandi и кариотип этой тетраплоидной рыбы, содержащий 90 хромосом.
Credit:
Nature (2026). DOI:
10.1038/s41586-026-10439-1 |
CC BY
Предполагается, что предки всех позвоночных претерпели полногеномную дупликацию, а впоследствии их геном вернулся к диплоидному состоянию. Однако ранние этапы этого процесса не описаны, поэтому неясно, как мог реализоваться потенциал, создавшийся из-за удвоения генетического материала — источника эволюционного разнообразия. Авторы статьи в Nature получили и проанализировали геномы двух видов карповых рыб подсемейства Schizothoracine, обитающих в высокогорных водоемах. Это сравнительно молодая таксономическая группа, еще не претерпевшая редиплоидизацию — все представители обладают как минимум тетраплоидным геномом. Ученые подтвердили, что предок Schizothoracine прошел одно событие тетраплоидизации перед расхождением на разные виды.
Подсемейство Schizothoracinae возникло от 10 до 30 миллионов лет назад во время резких геологических и климатических изменений на Тибетском плато. Оно включает более 100 аутополиплоидных (4–20n) видов из 11 родов, по фенотипическим адаптациям отнесенных к трем группам — «неспециализированной», «специализированной» и «высокоспециализированной». Авторы секвенировали геномы представителей всех 11 родов. Они получили также митохондриальные геномы и построили по ним филогенетическое древо, которое подтвердило ранее предполагавшиеся родственные связи и выделило неспециализированную, специализированную и высокоспециализированную группы Schizothoracinae.
Для сборки генома de novo авторы выбрали высокоспециализированный вид Schizopygopsis younghusbandi и секвенировали геномы трио «родители-потомок». Потомков секвенировали с помощью PacBio HiFi-прочтений с покрытием более 100×, Oxford Nanopore с покрытием более 50× и Hi-C с покрытием более 250×, родителей — методом коротких прочтений с покрытием 100× и более. Затем ученые сопоставили независимые сборки по данным длинных прочтений и скаффолд, полученный методом HiC-прочтений.
Окончательная сборка содержала 90 хромосом, что соответствовало наблюдаемому кариотипу. Авторы сравнили результат со стандартной стратегией анализа трио и собрали 45 материнских и 45 отцовских хромосом в геноме потомства, что подтвердило эффективность изначального подхода.
Хотя гаплоидный набор Schizothoracinae содержит 25 хромосом, сборка и анализ кариотипа S. younghusbandi выявили в общей сложности 90 вместо ожидаемых 100 (4n = 25 × 4). Это предполагает слияние хромосом либо их утрату после полиплоидизации. Сравнение с аутгруппой Onychostoma macrolepis выявило признаки слияния в пяти парах хромосом.
Чтобы подтвердить хромосомные слияния, авторы воспользовались флуоресцентной гибридизацией in situ (FISH). Они пришли к выводу, что исходные тетраплоидные группы хромосом разделились на слитые (f) и неслитые (uf) копии, которые развивались по разным эволюционным путям. Их обозначили как f (m), f (p), uf (m) и uf (p), где m и p указывают на материнское и отцовское происхождение соответственно.
Дальнейший анализ различных тетраплоидных генотипов (AAAA, AAAa, AAaa, Aaaa и aaaa) в пределах одного генома показал, что в претерпевших слияние (f) квартетах хромосом больше дисомных генотипов, чем в квартетах без слияния (uf), где, напротив, больше тетрасомных генотипах. При этом дисомные генотипы в основном располагались в участках слияния, то есть именно в них началась редиплоидизация.
Проанализировав частоты аллелей у S. younghusbandi и девяти других видов, авторы выявили максимальное обогащение дисомными генотипами на хромосомах 19 и 22 — по-видимому, они претерпели редиплоидизацию у общего предка. Также ученые обнаружили дисомное обогащение в шести других хромосомах (4, 8, 15, 16, 20 и 23) у высокоспециализированных видов Schizothoracinae.
Наконец, исследователи сравнили S. younghusbandi с представителем неспециализированной группы — Schizothorax curvilabiatus — имеющим 98 хромосом. В первую очередь они изучили хромосомы 19 и 22, поскольку, если редиплоидизация произошла у общего предка, то тогда же должны были дивергировать пары онологов (копий генов, возникших в результате полногеномной дупликации — это частный случай паралогичных генов). Анализ показал, что редиплоидизация этих хромосом, связанная с их слиянием, действительно началась у общего предка. Однако на момент расхождения его на различные виды она была неполной, а затем продолжалась в каждой линии независимо.
Таким образом, работа объясняет фундаментальный механизм, по которому претерпевшие полиплоидизацию виды начали возвращаться к исходному диплоидному состоянию. За переход от тетрасомного к дисомному наследованию отвечают отдельные слившиеся группы хромосом, авторы предполагают, что именно это инициировало повторную диплоидизацию у Schizothoracinae. Сейчас представители этого семейства переживают процесс возврата к диплоидному состоянию.
Источник
Chromosomal fusions trigger rediploidization of autopolyploid genomes. // Nature (2026). DOI:10.1038/s41586-026-10439-1
Вам будет интересно

 213
213
 0
0